図1
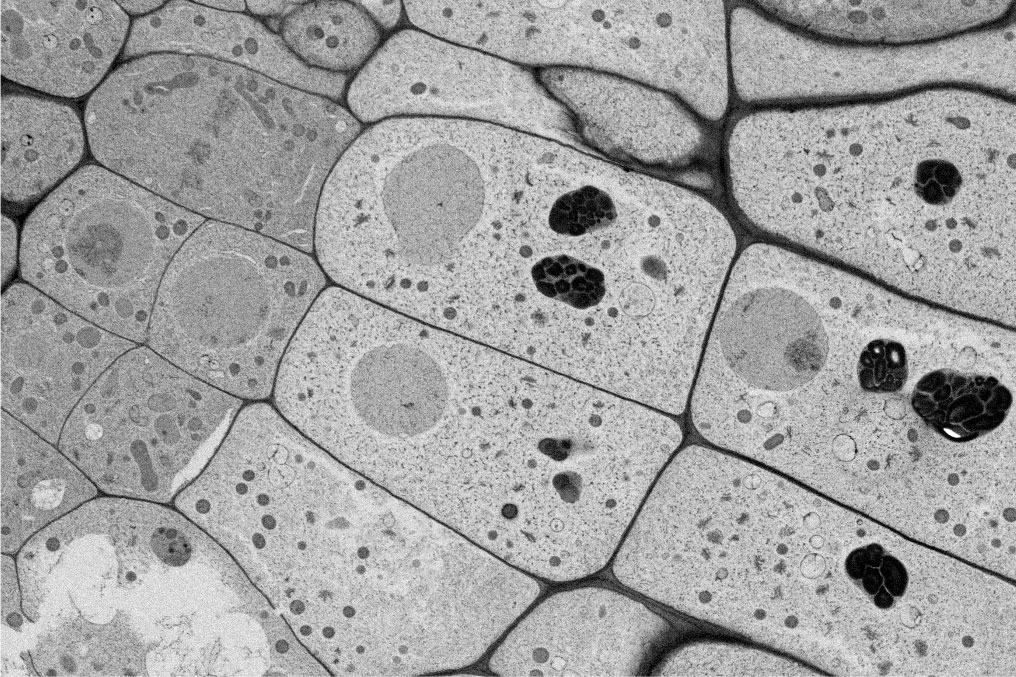
図1.細胞脱分化にともなう色素体の変化
たとえば傷害ストレスで誘導されるカルスは、由来する組織・細胞にかかわらずほぼ無色の細胞で成り立っています。カルス化の際には、色素体の変化が起きることも古くから知られています。葉緑体が発達した葉肉細胞からは、プロプラスチドやアミロプラストを有したカルス細胞が誘導されます。
Figure 1. Plastid reprogramming accompanied by cell dedifferentiation
The formation of callus consisting of colorless cells is induced by wounding, which triggers plastid reprogramming. Mesophyll cells with chloroplasts give rise to callus cells containing proplastids or amyloplasts.